
Artículos
Pastoreo Estratégico de Alto Impacto, una Herramienta para Optimizar el Crecimiento de los Pastizales en el Norte de Corrientes

Autores: D. B. Kurtz, F. Asch, M. Giese, C. Hülsebusch, M. C. Goldfarb and J. F. Casco
Traducción: D. D. Ybarra y D. B. Kurtz
Resumen
La principal fuente de alimentación para el ganado en la Argentina lo constituyen los pastizales. Muy a menudo en el noroeste de la provincia de Corrientes la biomasa muerta y seca en pie tiende a acumularse a partir del otoño y constituye una barrera para la utilización ganadera eficiente del forraje producido. En este trabajo se evaluaron los efectos del pastoreo de alto impacto (PAI) sobre la producción de biomasa en pastizales del noroeste de Corrientes, aplicado en diferentes épocas del año, aplicado para eliminar los excesos de biomasa muerta en pie. El PAI se realizó en forma mensual en potreros adyacentes a lo largo de un año y se evaluaron sus efectos hasta 1 año posterior a su aplicación. Se estudió la dinámica del crecimiento de la biomasa, la acumulación de biomasa verde y la acumulación de biomasa muerta en pie. En general el PAI mejora la relación entre la biomasa verde y la biomasa total. La época del año en que se aplica el PAI produce un efecto muy marcado en la dinámica de la tasa de crecimiento del pastizal. El PAI produce una fase de crecimiento activo durante el otoño, cuando en el testigo la biomasa verde disminuyó significativamente. Los mejores resultados en términos de reducción de la biomasa muerta en pie se lograron después del PAI en invierno, mejorando la relación biomasa verde – biomasa muerta. El PAI en otoño, sin embargo, reduce la disponibilidad de forraje y reduce la productividad del pastizal del año siguiente. El PAI realizado en tiempo y forma, constituye una herramienta estratégica, no sólo como un método alternativo para reducir la biomasa muerta en pie, sino también como un camino hacia la intensificación sostenible, proporcionando forraje verde a niveles iguales o incluso superiores a los alcanzados en pastoreo continuo tradicional.
*Artículo original en inglés publicado como: High impact grazing as a management tool to optimize biomass growth in northern Argentinean grassland – D. B. Kurtz, F. Asch, M. Giese, C. Hüsebusch, M. C. Goldfarb and J. F. Casco, Ecological Indicators, 63, 100-109 (2016).
Introducción:
Disturbios tales como las quemas y el pastoreo han transformado la estructura de los pastizales de la Argentina (Carnevali 1994). En el norte de la provincia de Corrientes, con fuerte tradición ganadera, la productividad primaria neta (PPN) de las gramíneas C4 es muy alta en verano y relativamente baja en invierno (Bernardis et al. 2005b; Martín et al. 2011; Royo Pallarés et al. 2005). Por lo tanto y a pesar de la alta producción primaria, la relativamente baja carga animal de estos pastizales (bajo uso ganadero tradicional), se ajusta a la relativamente baja disponibilidad del forraje invernal (Calvi et al. 2010). Gran parte de la biomasa no se consume y como consecuencia, la biomasa muerta y seca en pie se acumula en grandes áreas de pastizales en el noroeste de Corrientes (Kurtz et al. 2010).
Un considerable potencial de producción sub-utilizado
Por un lado, la biomasa muerta produce sombra que disminuye la captación de energía para la fotosíntesis, que a su vez disminuye la producción neta (Fidelis et al. 2013). Por otro lado, la biomasa muerta en pié no solo actúa como un elemento de disuasión al pastoreo (Balph and Malecheck, 1985; Moisey et al. 2006), sino que también reduce la ganancia de peso vivo de los herbívoros debido a la pobre palatabilidad y la baja calidad general de ese forraje (Mingo and Oesterheld, 2009). Debido a estas y otras razones, la producción animal en general en los pastizales del norte de Argentina es baja (Royo Pallarés et al. 2005). Según datos recientemente publicados, las tasas de ganancia de peso promedio del ganado se han mantenido constantes en los últimos 60 años en Corrientes (Calvi et al. 2010).
Métodos y momentos para eliminar el material muerto en pié
Existe una amplia gama de posibles tratamientos para reducir o eliminar el material muerto en pie. Por un lado está la eliminación mecánica, por ejemplo, haciendo uso de rolos con cuchillas, el desmalezado y el uso de arados (Adema et al. 2004). Sin embargo, el fuego es la práctica más frecuente y más fácil de usar en los pastizales y sabanas tropicales (Oesterheld y otros, 1999; Pausas et al 2013). A pesar de sus ventajas, tanto el fuego como las opciones mecánicas de control tienen sus desventajas, a veces la quema aumenta el riesgo de invasión de arbustivas (Fidelis et al 2013; Thomas, 2006. Dudinszky y Ghermandi, 2013) y/o puede provocar la pérdida de biodiversidad (Azpiroz et al 2012; Podgaiski et al 2014.). A su vez, las opciones mecánicas pueden producir la compactación del suelo (Hamza y Anderson, 2005; Jung et al 2010; Schrama et al 2013) y reducir la infiltración de agua (Chyba et al 2014). Recientemente y por ley, se ha limitado la quema tanto en la Argentina (Argentina, 2009), como en la Provincia de Corrientes (Corrientes, 2004), por lo que resulta importante la búsqueda de nuevas alternativas de eliminación del material muerto en pié que sean económica y ambientalmente posibles. En este sentido, el pastoreo de alto impacto (PAI) o «efecto manada» se propuso como opción de manejo para la restauración y mantenimiento de las funciones de los ecosistemas de pastizal (Savory 1983; 2005) y como un medio para mejorar la productividad de los pastizales (Savory y Parsons, 1980). El PAI en primer lugar elimina el sombreado ocasionado por la biomasa muerta produce y por el efecto mecánico de pisoteo produce una re-distribución – homogeneización del suelo y la incorporación de las excretas, por lo que mejora el ciclado de nutrientes (Briske et al. 2008; 2011; 2013; Dunne et al. 2011; Joseph et al. 2002). En algunos ecosistemas de pastizales se ha demostrado que el PAI incluso estimula el crecimiento y la calidad forrajera de la vegetación (McMillan et al 2011; Cromsigt y Olff, 2008; Hampson et al 2014; McNaughton, 1984).
Aunque el problema antes mencionado es largamente conocido, la información de los efectos del PAI sobre la dinámica del pastizal es sorprendentemente escasa y el uso del efecto manada ha generado una fuerte controversia en la comunidad científica (Briske et al, 2008, 2011, 2013; Dunne et al, 2011; Joseph et al, 2002). Son sólo unos pocos estudios los que analizaron los efectos del PAI en la producción de biomasa a largo plazo. Jacobo et al. (2000; 2006) encontraron que el pastoreo rotativo produce efectos positivos en la producción al reducir el material muerto en pie, mientras que Striker et al. (2011) encontraron que en praderas inundadas la proporción gramíneas se incrementa después de un PAI, mientras que la productividad primaria neta aérea no fue afectada significativamente.
La mayoría de los ecosistemas de pastizales se caracterizan por la estacionalidad, es por eso que la época en que se realiza el PAI (primavera, verano, otoño o invierno) probablemente afecte la dinámica del crecimiento de la biomasa durante los meses siguientes al PAI. Conocer la época ideal del PAI que produzcan cambios positivos considerables en la relación biomasa verde / biomasa muerta y la productividad de los pastizales, mejoraría finalmente la producción animal. No hemos encontrado evidencias que indique si el PAI constituye una práctica de manejo alternativa para el manejo de los pastizales del norte argentino, tanto para controlar la biomasa muerta como para promover la producción. Los resultados aquí presentados son relevantes para el desarrollo de estrategias de intensificación sustentable de uso de la tierra respetando la estabilidad del ambiente, a la vez que se incrementa la productividad pecuaria (Garnett et al. 2013).
Materiales y métodos:
Área de estudio
El estudio se llevó a cabo en la Estación Experimental Corrientes del INTA (27°40’01 »S, 58° 47’11»O), Departamento Empedrado, a 30 km al sur de la ciudad de Corrientes, Capital de la Provincia de Corrientes, Argentina. El sitio está a 69 metros sobre el nivel del mar y la pendiente general del terreno es de menos del 0,1%. La precipitación anual media es de unos 1300 mm, con una ligera estacionalidad de las lluvias, la mayor parte de la precipitación ocurre en el otoño (33% de marzo a mayo) y verano (30% de diciembre a febrero), algo menos en primavera (24% de septiembre a noviembre) y menor en el invierno (13% de junio a agosto). La temperatura media anual es de 21°C. La amplitud térmica anual oscila entre 25,6°C la media mensual en enero a 15,5°C la media mensual en julio.
La temperatura media durante el experimento fue similar a la de la temperatura media histórica. La precipitación durante el período experimental varió sólo ligeramente entre años, a partir de junio 2012 a mayo 2013, la precipitación total fue de 1.345 mm, y la evapotranspiración fue de 1.150 mm. Desde junio 2013 hasta mayo 2014, las precipitaciones fueron 1.233 mm y la evapotranspiración 1.107 mm (Fig. 1).
Los suelos en el sitio de instalación del ensayo son de textura franco-arenosa y pertenecen a la serie de Treviño (Argiudol Aquico, Escobar et al., 1996). Estos suelos se mantienen húmedos o muy húmedos durante la mayor parte del tiempo todos los años, sobre todo debido a la alta precipitación y a la capa de arcilla situada a unos 40-90 cm de profundidad (horizonte Bt). El pH varía de 5,6 a 6,0 en superficie, hasta a 7,0 a 7,4 en el Bt. La materia orgánica del suelo varía entre el 1,2 al 1,7% en la parte superior, a apenas 0,3% a 90 cm de profundidad (Escobar et al., 1996).
Vegetación dominante:
En los potreros donde se llevó a cabo el ensayo, las especies dominantes que forman matas son: la paja colorada (Andropogon lateralis Nees), la paja amarilla
(Sorghastrum setosum (Griseb.) Hitchc.) y Paspalum plicatulum Michx. Mientras que entre las matas se desarrollan otras especies de porte bajo, como el pasto horqueta (Paspalum notatum Flügge), Axonopus affinis Chase, Eleocharis nodulosa (Roth) Schult., E. viridans Kuk. ex. Osten. y Leersia hexandra Sw. Las leguminosas son bastante escasas, encontrándose a veces Desmodium incanum DC; y Vicia epetiolaris Burk (Vanni y Kurtz, 2005).
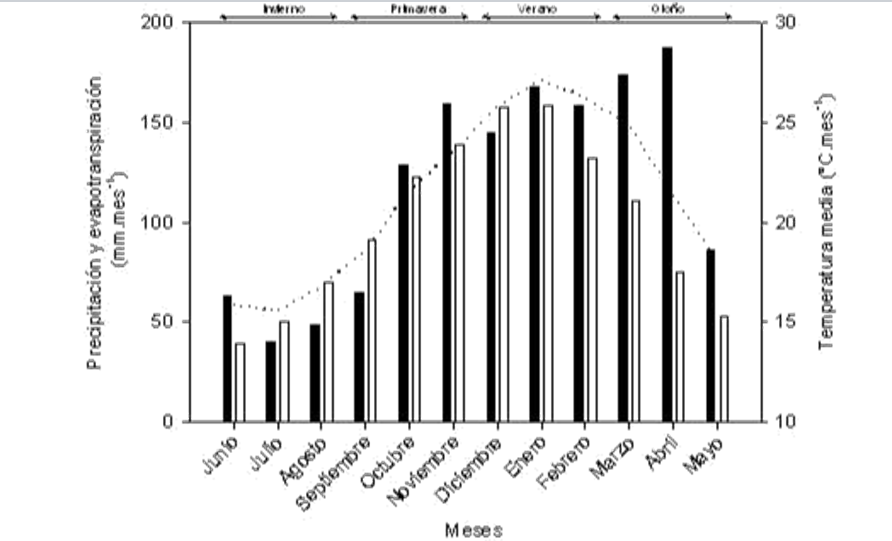
Figura 1. Patrones climáticos mensuales. Datos meteorológicos de INTA Corrientes, periodo 1968-1998 (parte superior derecha) y durante el experimento 2012- 2014 (abajo a la derecha). La línea punteada indica la temperatura media del aire (° C). Las barras blancas indican la evapotranspiración mensual de referencia y barras negras representan la precipitación mensual (mm).
El Experimento
El experimento se estableció en un pastizal natural, sobre una superficie de 24 hectáreas que forman parte de la Estación Experimental del INTA Corrientes. En el pasado el área había sido manejada tradicionalmente con pastoreo continuo y con una intensidad de carga de 0,5 equivalente vaca hectárea-1año-1. Cuatro potreros adyacentes de 6 hectáreas cada uno se separaron con alambrados eléctricos permanentes. Tres potreros se utilizaron como repeticiones para el experimento de PAI (R1 – R3) y el cuarto se definió como testigo sin PAI. Todos los tratados y el testigo, permanecieron bajo pastoreo continuo con una carga de 0,5 equivalente vaca hectárea-1año-1.
El tratamiento de PAI siguió una secuencia mensual en lotes adyacentes. Cada repetición (R1, R2 y R3) se dividió en 12 lotes de 0,5 hectáreas cada una, donde se realizó el PAI mes a mes en lotes adyacentes. El experimento comenzó en julio de 2012, cuando el primer lote (de 50 m de ancho por 100 m de longitud) fue cerrado con alambres eléctricos móviles / temporales y sometido a tres días de PAI. Para ello se utilizó un lote mixto de 75 animales tipo Bradford, Hereford y Brahman, representando una carga instantánea de 150 animales ha-1 (aproximadamente 30000 kg de biomasa animal ha-1). Durante el primer día los animales pastaron libremente, a partir de segundo día la manada fue inducida a pisotear el pasto, conduciendo a los animales dentro del lote hasta que toda la vegetación fue aplastada o incorporada al suelo y garantizar así un impacto lo más homogéneo posible. Después de cada PAI, los alambres del lote se retiraban y la manada conducida a las dos repeticiones restantes, así se hizo sucesivamente mes a mes, en lotes consecutivos. Como se dijo anteriormente, los cuatro potreros (repeticiones y testigo) se pastorearon continuamente durante todo el experimento con 3 vaquillas cada uno, emulando la carga animal promedio en la provincia de Corrientes, de 0,5 equivalente vaca hectárea-1año-1 (Calvi 2010; Kurtz y Ligier, 2007), estas vaquillas se pesaron al inicio y al final del ensayo.
Producción vegetal del pastizal
El crecimiento mensual del pasto se midió utilizando dos jaulas protectoras (de 1m2) por lote, las jaulas se colocaban en el m2 recién cortado y se cosechaban mes a mes. Para calcular la disponibilidad de pasto por fuera de las jaulas de exclusión, también se cosechó mensualmente la biomasa, pero en este caso se clasificaron las muestras en material verde y muerto. Todo el material cosechado se pesó en húmedo y luego se secó en estufa a 75° C por 72 hs para determinar la materia seca.
Resultados:
El PAI no produjo un efecto negativo sobre el rebrote mensual de los pastos comparado con el testigo (Fig. 2). Tampoco hubo interacción entre la época de corte y el PAI (p = 0,2898). Sin embargo, la época de corte influyó significativamente en el rebrote de los pastos (p <0,0001); es decir, en el invierno se produjo el menor rebrote mensual (30 g m-2), mientras que las otras épocas el crecimiento se duplicó de manera significativa, siendo en el verano de 73 g.m-2, en el otoño de 64 g.m-2 y en la primavera de 60 g.m-2.
La Figura 3. muestra la dinámica de la biomasa total (la suma de la biomasa muerta en pié y la biomasa verde), tanto del pastizal tratado con PAI (según el momento del impacto, invierno, primavera, verano, otoño), como también la dinámica del pastizal en el testigo. Nuestros cálculos indican que, en promedio, el efecto inmediato del PAI produce una reducción de la biomasa total en un 95% (+ -1%).
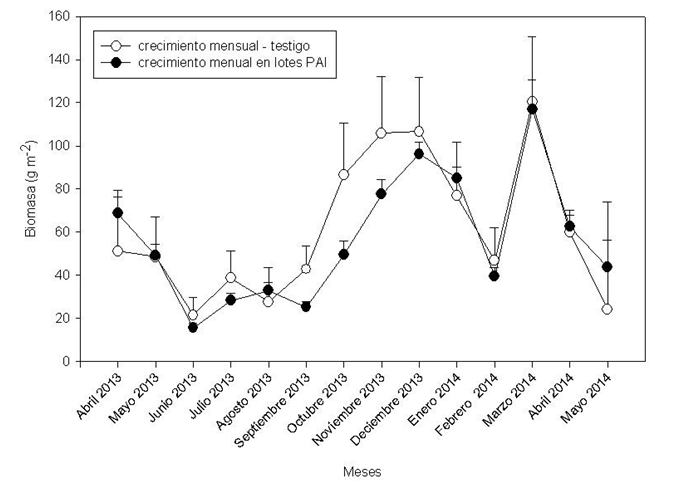
Figura 2. Dinámica de los pastizales, crecimiento mensual en el testigo y en los lotes de pastoreo de alto impacto (PAI), medido en jaulas de exclusión de pastoreo. Las variables se expresan en g.m-2. Las barras de error indican el error estándar de las medias (p <0,05).
El PAI reduce la biomasa total desde 1970 g m-1 en primavera, 1680 g m- 1 en verano, 1770 g m-1 en otoño y 2370 g m-1 en invierno, hasta aproximadamente 100 g m-1, de biomasa medida enseguida después del PAI. Durante todo el período experimental la biomasa total fue significativamente más baja según las diferentes épocas de aplicación del tratamiento de PAI (Fig. 3). La dinámica del pastizal en el testigo sigue un patrón estacional con un pico máximo de acumulación de biomasa entre noviembre y diciembre y otro mínimo entre abril y agosto. La biomasa total estuvo siempre por encima de 1000 g de materia seca por m-2. Por otro lado el pastizal sometido a PAI presentó valores de biomasa total entre 200 – 800 g m-2.
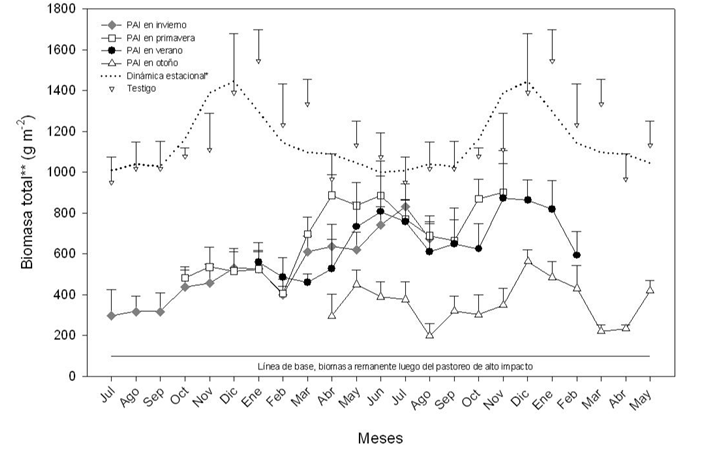
Figura 3. Evolución de la biomasa total después del pastoreo de alto impacto en invierno, primavera, verano y otoño, en comparación con la biomasa total cosechada en el testigo. Para una mejor comprensión de la dinámica de la biomasa en el testigo, ésta fue ajustada visualmente (línea de puntos). **Biomasa total = biomasa verde + biomasa muerta y seca en pié.
Además, en la figura 3 se puede observar que tanto en el testigo, como en los lotes de PAI se desarrollaron fases de activo crecimiento desde septiembre a enero (primavera y verano). A partir de entonces la biomasa total del testigo se redujeron en aproximadamente un 40% en el período de febrero a agosto (otoño a invierno). Por el contrario, en los lotes de PAI e independientemente del momento del impacto, se produjo una prolongación del período de crecimiento en el otoño (de febrero a junio), donde la biomasa se incrementó hasta 850 g m-2, la única excepción se dio con PAI realizado en otoño. La biomasa en el testigo disminuyó o se estancó en aproximadamente 1000 g m-2.
Época de aplicación del pastoreo de alto impacto (PAI)
La figura 4. Muestra la dinámica del pastizal después de PAI en primavera, verano, otoño e invierno, las dos curvas representan la biomasa total y la biomasa muerta, la diferencia entre ambas es la cantidad de biomasa verde en los lotes de PAI y testigo. El PAI en invierno produjo dos fases de crecimiento, un fuerte incremento de biomasa en la primavera y otro en el otoño (Fig. 4a). En contraste, en el testigo se produjo una sola fase de crecimiento activo en la primavera. Después del PAI en invierno, la acumulación total de biomasa en primavera fue más rápida en comparación con el testigo, expresada por la pendiente de la regresión que representa la tasa de crecimiento (pendiente b = 258 g m-2 vs. 196 g m-2 mes-1, Fig. 4a). La segunda fase de crecimiento del PAI de invierno incrementó la biomasa en torno a 500 g m-2. Por otro lado, en el testigo se produjo una disminución en la cantidad de materia seca en torno a los 300-400 g m-2. El PAI en primavera produjo una fase de crecimiento activo que se prolonga hasta otoño con un aumento de la biomasa (de hasta 1000 g m-2) evidente hasta siete meses después del impacto (Fig. 4b). Durante el mismo tiempo, el testigo mostró una disminución de la biomasa de entre 1500-1000 g m-2. A través del año, la mayor parte de la biomasa en el testigo tenía entre 62 a 84% de biomasa muerta seca en pié, en comparación con el 34 al 74% en los lotes de PAI. Por otra parte, la cantidad de biomasa verde no presentó valores significativamente diferentes (277 g m-2 vs. 252 g m-2 en el testigo y en los lotes de PAI, respectivamente). El PAI aplicado en el verano también promovió el crecimiento adicional de los pastos, la primera fase de crecimiento se produjo durante el otoño (esta fase estuvo ausente en el testigo y el pastizal mostró inclusive una tendencia negativa de acumulación de biomasa) y una segunda fase en la primavera. La fase de crecimiento de otoño dio lugar a un fuerte aumento de la biomasa total disponible (b = 137,1 g m-2 mes), que alcanzó un máximo de alrededor de 800 g m-2 (Fig. 4c). La segunda fase de crecimiento, en la primavera, comenzó en septiembre y se dio tanto en el pastizal tratado con PAI, como en el testigo. El PAI de otoño no provocó un segundo crecimiento activo de la biomasa en el año, pero resultó en una fase de crecimiento prolongada de septiembre a marzo, de manera similar al testigo. Durante este período, la biomasa total acumulada fue de aproximadamente 1000 a 1400 g m-2 en el testigo y de aproximadamente 300 a 700 g m-2 en las sub-parcelas sometidas al PAI en otoño (Fig. 4d). Independientemente de la época de aplicación del PAI, la cantidad absoluta de biomasa verde del pastizal fue casi siempre similar a la cantidad de biomasa verde en el testigo sin tratar. Hay que recordar que debido a la mayor cantidad de biomasa muerta seca en pié en el testigo, la proporción de biomasa verde fue siempre mayor en el pastizal sometido al PAI, al menos a lo largo de un año después del PAI (Fig. 5).
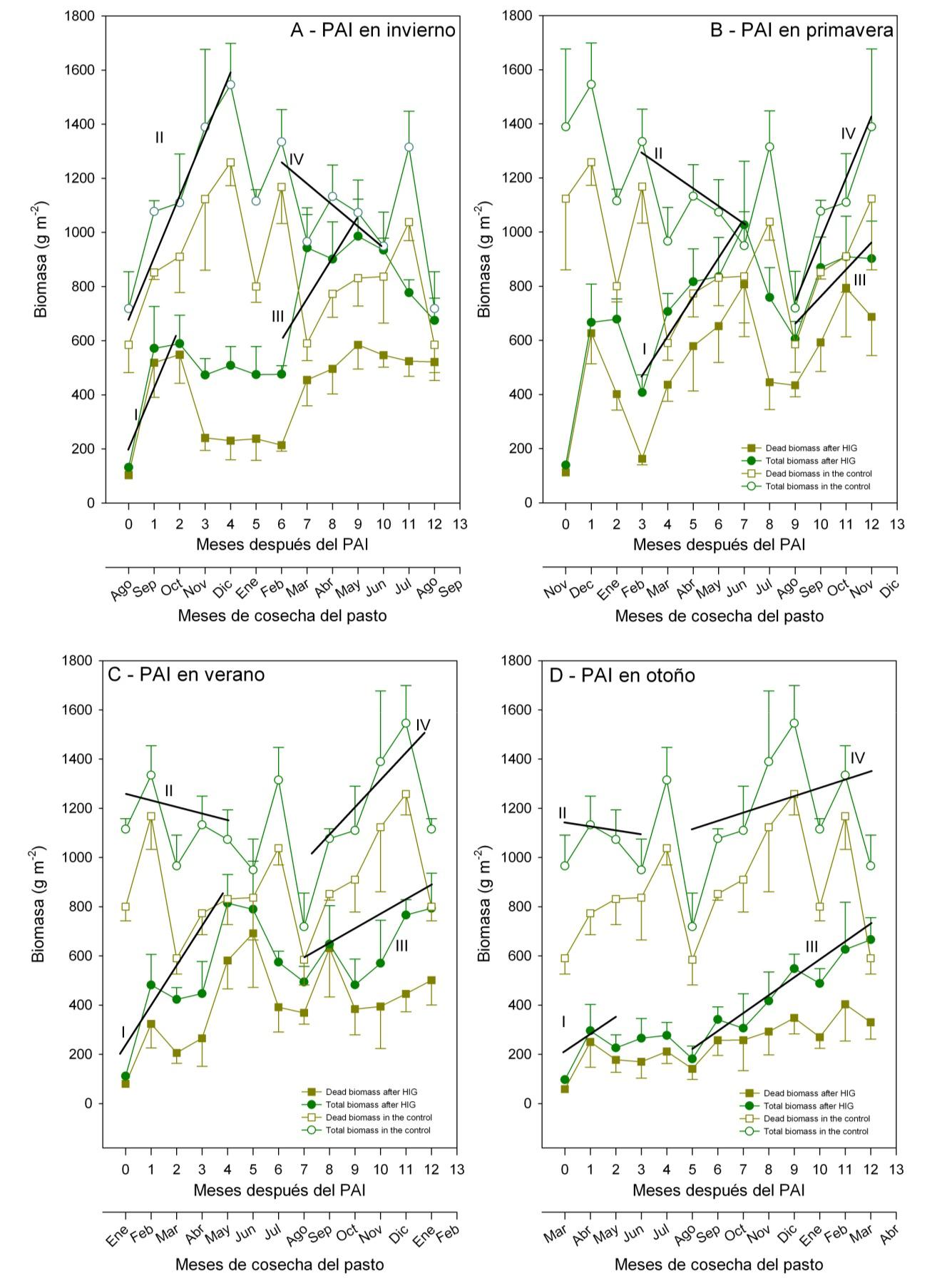
Figura 4. Dinámica de la biomasa del pastizal después de pastoreo de alto impacto (PAI) aplicado en las cuatro diferentes estaciones del año (primavera, verano, otoño e invierno). Ejemplos mostrados en esta figura: mes de agosto, PAI en invierno (A), para el mes de noviembre PAI primavera (B), para el mes de enero PAI en verano (C) y para el mes de marzo PAI en otoño (D).
La diferencia entre ambas curvas significa la biomasa verde. Para cada época de aplicación del PAI las regresiones se calcularon considerando los valores de biomasa total disponible, desde un mes después del PAI (mínima biomasa) y el momento en que se alcanza la máxima biomasa total disponible; similarmente en el testigo, y a efectos de hacer la comparación, la regresión se calculó teniendo en cuenta la biomasa total disponible durante ese mismo período de tiempo. La tasa de acumulación de biomasa fue muy distinta dependiendo de la época en que se aplicó el PAI, AII. y = x + 258,4 152,3 (r2 = 0,775); AII. y = x + 196,5 578,4 (r2 = 0,954); AIII. y = x + 148,9 603,5 (r2 = 0,661); AIV. y = x + -66,2 1.289,9 (r 2 = 0,452). BI. y = x + 92,9 681,9 (r2 = 0,686); BII. y = x + 196,5 484,9 (r2 = 0,902); BIII. HIG, y = x + 136,9 603,5 (r2 = 0,661); BIV. y = x + – 66,2 1.289,9 (r2 = 0,452). CI. y = x + 55,4 487,1 (r 2 = 0,6); CII. y = x + 104,8 792,6 (r2 = 0,472); CIII. y = x + 137,1 181,3 (r2 = 0,755); CIV. y = -28.5 x + 1209.9 (r2 = 0,113). DI. y = x + 88,9 504,6 (r 2 = 0,908); DII. y = x + 37,9 986,2 (r2 = 0,1273); DIII. y = x + 65,4 218,1 (r2 = 0,924); DIV. y = -10.9 x + 1057.6 (r2 = 0,026).
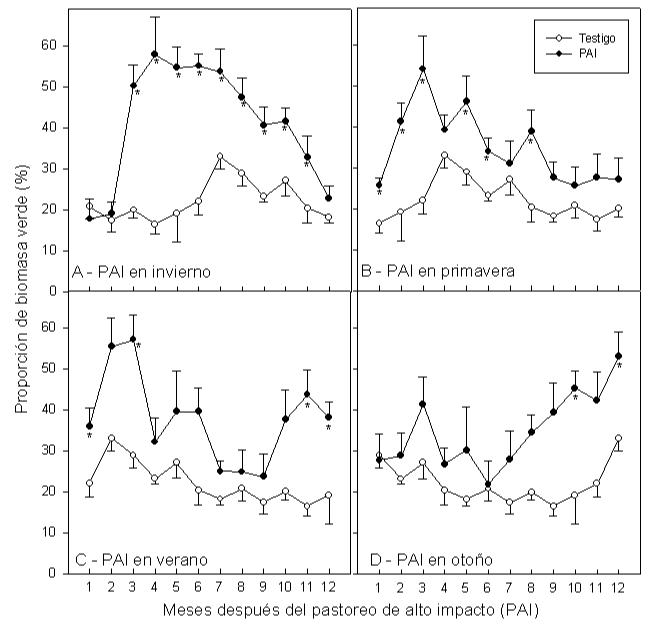
Figura 5. Proporción de biomasa verde en relación al tiempo transcurrido desde el PAI, aplicado en las cuatro diferentes estaciones. A modo de ejemplo se muestran los meses de: Agosto para el PAI en invierno (A), noviembre para el PAI en primavera (B), enero par el PAI en verano (C) y marzo para el PAI en otoño (D). Las barras de error indican el error estándar de las medias (p <0,05).
En el testigo, la proporción de biomasa verde fue mayor durante el verano, con máximos de alrededor del 30% de la biomasa total; sin embargo, en la mayor parte del año, la proporción de biomasa verde estuvo entre el 20 y el 25%. Por otro lado, en el pastizal sometido al PAI la proporción de biomasa verde alcanzó valores máximos una o dos veces al año, dependiendo de la época en que se efectuó el PAI, con proporciones de hasta el 60% de biomasa verde (Fig. 5). A lo largo del año, la proporción de biomasa verde en el pastizal sometido al PAI fue en promedio alrededor de 20% más alta que en el testigo. Se deduce claramente que en los lotes de PAI la biomasa disponible fue por lo tanto, más palatable y más fácilmente accesible a las vaquillas que en el testigo.
Discusión
El efecto del pastoreo de alto impacto en la dinámica del pastizal
El rebrote mensual de los pastos mostró un claro patrón estacional, típico de los pastizales dominados por gramíneas C 4, donde las bajas tasas de crecimiento coinciden con los períodos de baja temperatura y baja radiación (Knapp y Medina, 1999; Martín et al. 2011; Ötztürk et al. 1981; Royo Pallarés et al. 2005). El rebrote fue apenas un 8% más alto (no significativo) en el testigo (857 g m-2) en comparación las parcelas sometidas al PAI (791 g m-2). En este sentido el PAI no produjo un crecimiento compensatorio como fuera sugerido por McNaughton (1979; 1983), aunque tampoco produjo una reducción de la productividad tras el PAI. Las tasas de crecimiento mensuales fueron similares en los lotes sometidos al PAI comparados con el testigo, lo que indica que el pastizal resiste las perturbaciones producidas por el PAI. Aparentemente el PAI no logra acelerar la mineralización de nutrientes (por la deposición de adicional heces) ni daña los meristemas intercalares y apicales, por lo que en líneas generales, se recupera rápidamente (Heckathorn et al. 1999).
Por otra parte, el PAI redujo significativamente la biomasa total y la biomasa muerta seca en pie. Se confirma que los efectos sobre la dinámica del pastizal dependen en gran medida de estación del año en que se aplica el PAI (McNaughton, 1983). El PAI derivó en un patrón de crecimiento anti-cíclico en comparación con el testigo, con una fase de crecimiento activo durante el otoño cuando la biomasa en el testigo disminuye hacia el final del otoño (debido a la descomposición de la biomasa), mientras que el activo crecimiento luego del PAI fue gracias a la eliminación del material senescente.
En el pastizal manejado tradicionalmente (testigo) hay crecimiento con acumulación de biomasa durante la primavera y el verano, luego la biomasa empieza a decaer y disminuye a finales de otoño hasta finales del invierno en agosto. La tasa negativa de la acumulación de biomasa está directamente relacionada con las condiciones climáticas, en particular a las bajas temperaturas (Long, 1999), pero también, asumimos que es probablemente debido al sombreado producido por la misma biomasa acumulada. Está comprobado que un dosel abierto, que permita una alta la intercepción de luz es esencial para producir altas tasas de fotosíntesis en las plantas C4 (Heckathorn et al. 1999; McMillan et al. 2011; Ötztürk et al. 1981). Como consecuencia se produce una alta producción de biomasa (Heckathorn et al. 1999; Pearcy et al. 1981). En el testigo, durante el otoño y el invierno se produce una combinación de altas cantidades de biomasa que produce sombra a las hojas inferiores y temperaturas más bajas. La penetración de luz debió haber sido mayor donde se produjo el PAI, demostrado por el crecimiento activo observado en otoño en los lotes sometidos a PAI, allí la relación entre biomasa verde y seca en pié fue mayor (Fig. 2).
En comparación con el PAI de invierno, verano o primavera (donde la acumulación de biomasa total fue entre ~ 400 a 800 g m-2), el PAI de otoño produjo excepcionalmente baja biomasa total (~ 200 a 600 g m-2) (Fig. 4d). Dos efectos principales pueden haber sido la causa de esto. Por un lado, la disminución de las temperaturas hacia el invierno al inducir a las plantas C4 a asignar recursos a los órganos subterráneos, es muy probable que el PAI en otoño haya impedido la asignación de los fotosintatos a las raíces (Knapp y Medina, 1999). Por lo tanto, el PAI en otoño, al destruir la biomasa interfirió con este proceso lo que se tradujo en un bajo crecimiento posterior. Además el PAI en otoño, con suelo saturado con agua pudo haber sido más dañino (Striker et al. 2011). La alta precipitación y la baja evapotranspiración durante el otoño de hecho resultaron en el encharcamiento temporario del suelo, por lo que el pisoteo en esas condiciones es probable que haya provocado lesiones y daños a las raíces y tallos (Dunne et al. 2011; Striker et al. 2006), lo que se tradujo en la reducción del crecimiento durante la primavera y hasta el verano siguientes. Es evidente que en el mediano término, los pastizales son más sensibles al PAI en otoño, cuando los suelos están normalmente saturados, tal vez si se hubiera aplicado de una manera menos perjudicial en esta época del año es probable que los daños fueran considerablemente menores.
Proporción de biomasa verde
En general, el pastizal del testigo ofreció enormes cantidades de material verde mezclado con material muerto seco en pié (entre 800 a 1.600 g m-2 de biomasa total) que lógicamente produce la disuasión al pastoreo (Balph y Malecheck, 1985; Moisey et al. 2006). La proporción de material verde en el testigo apenas llegó al
30% en otoño y en promedio en todo el año fue de alrededor de 22%. Por el contrario, la proporción de biomasa verde fue mucho mayor en los lotes donde se aplicó el PAI. Por ejemplo, promedió entre el 38 y el 42% después del PAI en invierno y en verano respectivamente (Fig. 5). Por otra parte, parece que debido a
la eliminación del material muerto seco en pié y la eliminación del sombreado también se prolongó la longevidad de las hojas (McNaughton, 1983), como se refleja en el mayor porcentaje de biomasa verde en los lotes sometidas a PAI comparadas con el testigo (Fig. 5). Si bien el PAI reduce la biomasa en torno al 95%; la estacionalidad (por ejemplo condiciones más húmedas o más secas) condicionan los efectos del PAI. En condiciones de suelos saturados y/o con agua en superficie, se produce una mayor incorporación de biomasa en el suelo.
Por último, en general y si bien la disponibilidad de forraje fue menor en los lotes sometidas al PAI en comparación con el testigo, después del PAI el pastizal mantuvo biomasa más verde y por ende más palatable durante todo el año comparado al testigo (Fig. 5). Independientemente de cuando se realizó el PAI, la biomasa de los pastizales se rejuveneció (McNaughton, 1983). Por otra parte, los resultados del presente estudio sugieren que se mejoró el acceso al pasto, debido a la eliminación del 95% del material muerto seco en pié. La proporción de biomasa verde (relación entre biomasa verde / biomasa muerta seca en pié) es un indicador de los efectos positivos del PAI y si bien, la cantidad de biomasa verde producida fue menor cuando el PAI se aplicó en verano u otoño en comparación con el impacto de invierno o primavera, los efectos positivos para el período de invierno y primavera (el período más difícil para la alimentación animal) cobran mayor relevancia para la productividad general del sistema. El PAI en cualquier época del año aumentó la relación verde/muerto, consecuentemente mejoró la captura de energía lumínica del pasto durante los períodos de invierno y principios de primavera, cuando el crecimiento normalmente es limitado por la materia muerta seca en pié.
Implicancias para el manejo de pastizales y la producción de carne
Por efecto del PAI la biomasa total se redujo, pero la cantidad de biomasa palatable y verde fue suficiente para mantener una carga animal de 0,5 EV a lo largo del año. A modo de ejemplo, durante los tres primeros meses después del PAI de invierno el pastizal tuvo suficiente biomasa verde (~ 170 kg ha-1) (que es similar a la carga media en la Provincia – considerando un consumo diario teórico de 12 kg de materia seca o el 3% del peso vivo de una vaca de 400 kg). Por otro lado, después de los PAI en primavera, verano u otoño, la biomasa verde disponible fue de entre 2 y 6 veces mayor a lo necesario para esa carga animal. Mientras que en el testigo, si bien durante todo el año la biomasa verde fue entre 4 y 10 veces mayor a los potreros donde se aplicó el PAI, la biomasa verde potencialmente consumible por el animal fue difícilmente accesible, debido al enorme volumen de material muerto y el efecto disuasorio que éste produce al pastoreo. Es muy probable que el pastoreo haya sido menos eficaz en el testigo ya que las vaquillas probablemente pasaron más tiempo y gastaron más energía en busca de forraje que consumiéndolo (Abdel-Magid et al 1987; Heckathorn et al., 1999).
Al principio del ensayo todas las vaquillas tenían el mismo peso vivo (232,8 kg, desvío estándar = 18,3 kg), tanto las que pastorearon los lotes bajo PAI como en el testigo. Al ser pesadas al finalizar el ensayo, las vaquillas del testigo pesaban 282,3 kg (desvío estándar = 19,1 kg), mientras que las que estaban en los lotes de PAI ganaron un 30% en peso vivo (400,9 kg, desvío estándar = 86,7, Fig. 6).
Nuestros resultados muestran claramente que el peso de las vaquillas aumentó significativamente en los pastizales sometidos al PAI comparado con el testigo.
Por otro lado, en los potreros de PAI, la combinación de áreas de escasa biomasa de alta calidad y áreas de gran volumen y de menor calidad, hayan contribuido a mejorar la utilización del pasto por parte de las vaquillas (Hempson et al. 2014).
Esto pudo haber contribuido a incrementar la ganancia de peso vivo de las vaquillas de los potreros de PAI, que fue superior a la de las vaquillas del testigo, donde pastorearon en un “mar” de material muerto. Las razones son especulativas, pero los resultados sugieren una mejor disponibilidad debido a las menores proporciones de material muerto en pié como resultado del PAI, o un mejoramiento de la calidad nutritiva de los pastos o el efecto combinado de ambos factores. Solamente el análisis de la calidad del forraje podrá revelar si el PAI mejoró el contenido nutricional de los pastos o no.
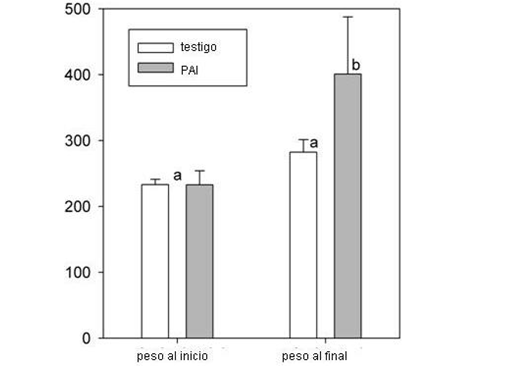
Figura 6. Peso vivo (kg) de las vaquillas al principio y al final del período evaluado (2013 a 2014), en el testigo y en los tratamientos de PAI. La figura muestra los pesos medios y las barras verticales indican la desviación estándar. Medias con la misma letra no son significativamente diferentes (p> 0,05).
Nuestros resultados sugieren que el PAI aplicado a finales del invierno es el más beneficioso para mejorar la disponibilidad de los pastos e incrementar la relación biomasa verde / biomasa muerta, prolongando el crecimiento. El PAI durante el otoño, sin embargo, puede I) reducir significativamente la disponibilidad de forraje durante el invierno y II) poner en peligro la productividad del año siguiente debido muy probablemente a la destrucción de las raíces por la pezuña del ganado, cuando el PAI se realiza sobre suelos saturados de agua. Somos conscientes de que son necesarios más estudios para recomendar el PAI como una herramienta de manejo del pastizal. Se necesitan más ensayos para entender mejor las interacciones planta-animal, para poder así utilizar este mecanismo de perturbación “casi natural” aprovechando así al máximo sus beneficios (Cromsigt y Olff, 2008; Hempson et al 2014; McNaughton, 1984) y aumentar así la eficiencia de uso de los recursos naturales y aumentar la productividad de los pastizales.
Conclusiones:
Proporcionamos una alternativa de manejo del material muerto seco en pié que contribuye a mejorar el consumo por parte del ganado. El efecto del PAI de reducir la biomasa muerta se prolonga durante varios meses, mejorando la proporción de biomasa verde del pastizal. Es fundamental considerar el momento de aplicación del PAI, considerando la dinámica estacional del pastizal natural (Figura 7). Los mejores resultados en términos de reducción de la biomasa muerta en pie y mejoramiento de las relaciones de biomasa verde / biomasa muerta se logran con PAI en invierno. El PAI en otoño, sin embargo, reduce la disponibilidad de forraje y la productividad de los pastizales del año siguiente al PAI. Independientemente de la estación en que se aplica el PAI se produce una fase de crecimiento prolongado que dura, por lo menos, hasta el otoño siguiente al PAI. Este crecimiento prolongado debe ser explotado por el potencial que supone al mejorar la disponibilidad de forraje para el ganado al comienzo del invierno. Nuestros resultados contribuyen a una mejor comprensión de los efectos de esta perturbación sobre el pastizal, con el potencial de ser utilizado para una mejor utilización de los pastizales correntinos. El PAI podría ser una alternativa valiosa, no sólo por ser un método alternativo de reducción de la biomasa muerta, sino también pensando hacia una intensificación sustentable que proporciona forraje verde y palatable, a niveles iguales o incluso superiores a los alcanzados bajo pastoreo tradicional continuo.
Figura 7. Eco-grama ilustrativo de los principios del PAI versus el manejo tradicional del pastizal.
Agradecimientos
Este trabajo fue financiado en forma conjunta por el Instituto de Ciencias Agropecuarias Tropicales (Institute of Agricultural Sciences in the Tropics -Hans-Ruthenberg-Institute) de la Universidad de Hohenheim, el INTA-Corrientes a través del PRET Noroeste y el servicio Alemán de Intercambio Académico (DAAD) con fondos del Ministerio Federal de Cooperación Económica y Desarrollo de la República Federal Alemana (BMZ) y la Fundación FIAT PANIS. La investigación tuvo además el aporte de una beca del Centro de Seguridad Alimentaria (FSC) de la Universidad de Hohenheim, parte del programa «exceed” del DAAD. Los conceptos, ideas y el marco lógico de este trabajo fueron desarrollados en cooperación con investigadores de la red GrassNet del DAAD.
Los autores agradecen la colaboración de Cristina Goldfarb, José Casco, Juan José Verdoljak, Joaquín Flores, Diego Ybarra y al staff de servicios generales del INTA Corrientes, específicamente al Sr. Jorge Godoy, Lucio Almirón, Oscar Ramón Arce, Leocadio Acosta, Aldo Luis Silva Bizarro, Juan Aranda, Gregorio Santiago Prieto, Ramón Alfonso Ruiz Díaz y Eugenio Mendoza. También y de manera muy especial gracias a Francisco Nuñez. Raúl Aranda, Oscar Gerónimo Quiros Villalba, Marcelino Retamozo, Gustavo Antonio Prieto, Ismael Palacios, Rodolfo Alberto Fonteina, Marcos Colman y Marcelino Raúl Fernández. Gracias a los integrantes del grupo Recursos Naturales Héctor Daniel Ligier, Rosa Lencinas, Marcelo Garay, Raúl Barrios, Humberto Matteio, Osvaldo Vallejos, Ruth Perucca, Sandra Perucca, Cristina Sanabria, Soledad Rey Montoya, Shinyi Hinomata Kaneoya, Julia Bernardi, Reimunda Falcón y Mercedes Falcón. Inmensas gracias a Leon Wurtz, Eugen Jedig, Saskia Windisch, Rabea Mächtel, Karen Kauselmann, Alberto Escobar, Liliana Escobar, Guillermo Frank and Juan José García Alevras por su ayuda.















-

 Artículos1 año atrás
Artículos1 año atrásGuía de Manejo de Semen Bovino
-

 Noticias Nacionales11 meses atrás
Noticias Nacionales11 meses atrásConsumo de bebidas: ¿cuál es la categoría que más crece en 2025 y por qué?
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásGloria lanza programa “Productividad Ganadero” a favor de la producción de leche en el Perú
-

 cursos on line1 año atrás
cursos on line1 año atrásCurso Virtual: Actualización en Sanidad Bovina – Control, Inmunología y Manejo de Vacunas
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásAgrovet Market lanza nueva línea de antibióticos en suspensión para el tratamiento de la mastitis
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásGloria: la primera marca peruana en el Kantar Creative Effectiveness Awards 2025
-

 Cursos A Distancia8 meses atrás
Cursos A Distancia8 meses atrásCurso Virtual: Formulación de Raciones para Ganado Lechero con software especializado NASEM
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásGloria amplía “PRO”: portafolio extendido con proteínas enfocadas en la energía