
Artículos
Anatomía y Fisiología de la Reproducción Bovina
Mel DeJarnette, Especialista en reproducción
Dr. Ray Nebel, Especialista en Reproducción
Todo programa exitoso de inseminación artificial está basado en un amplio conocimiento de la anatomía y fisiología reproductiva de los bovinos. Antes de intentar inseminar una vaca, debes hacerte una gráfica mental de los órganos que componen el aparato reproductor femenino. Para poder entender el porqué un animal exhibe síntomas de celo, cuando se debe inseminar, y como se desarrolla la preñez, se debe tener un claro entendimiento de los mecanismos hormonales que controlan el ciclo estral en las vacas.
ANATOMIA
Primero, demos una mirada a las partes que componen el parto reproductor bovino (Figura 1). Hay dos Ovarios, dos Oviductos, dos Cuernos Uterinos, un Utero, la Cervix, la Vagina y la Vulva. La Vejiga está ubicada debajo del aparato reproductor, y está conectada a la apertura uretral en la base de la Vagina.
El Recto está ubicado encima del aparato reproductor.
La Vulva es la apertura externa del aparato reproductor. Ella tiene tres funciones principales: dejar pasar la orina, abrirse para permitir la cópula y sirve como parte del canal de parto.
Incluidos en la estructura vulvar están los Labios y la Clítoris.
Los Labios de la Vulva están ubicados a los lados de la apertura vulvar, y tienen aspecto seco y arrugado cuando la vaca no está en celo. En la medida que el animal se acerque al celo, la Vulva empezará a hincharse y tomará una apariencia rojiza y húmeda.
La Vagina, que tiene como seis pulgadas de largo, se extiende desde la apertura uretral hasta la Cervix. Durante la monta natural, el semen es depositado en la porción anterior de la Vagina.
La Vagina también sirve como parte del canal de parto al momento del parto.
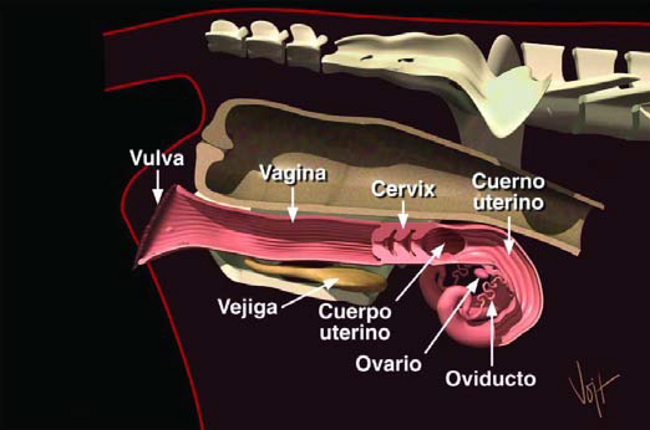
 figura 1. Vistalateral sistema reproductor femenino.
figura 1. Vistalateral sistema reproductor femenino.
«Para poder entender el porqué un animal exhibe síntomas de celo, cuando se debe inseminar, y como se desarrolla la preñez, se debe tener un claro entendimiento de los mecanismos hormonales que controlan el ciclo estral en las vacas.»
La Cervix es un órgano de paredes gruesas, que establece la conección entre la Vagina y el Utero de tejido conectivo denso y músculos, y será nuestra referencia al inseminar una vaca. La entrada a la Cervix está proyectada hacia la Vulva en forma de cono. Esto forma un círculo ciego de 360º que rodea completamente la entrada a la cervix. Esta base ciega del cono es conocida como Fornix. El interior de la Cervix contiene tres o cuatro Anillos, a veces llamados pliegues. Este diseño le facilita a la Cervix ejercer su función principal, que es la de proteger el Utero del medio ambiente exterior. La Cervix se abre hacia adelante al Cuerpo Uterino. Como de una pulgada de largo, el Cuerpo Uterino sirve de conección entre los dos Cuernos Uterinos y la Cervix. El Cuerpo Uterino es el sitio donde se debe depositar el semen durante la Inseminación Artificial.
A partir del Cuerpo Uterino, el tracto reproductor se divide y todos los órganos vienen en pares (Figura 3). Los dos Cuernos


 Figura 3. El Utero se divide en dos Cuernos Uterinos.
Figura 3. El Utero se divide en dos Cuernos Uterinos.
Uterinos están formados por tres capas musculares y una intricada red de vasos sanguíneos. La función principal del Utero es proveer el ambiente óptimo para el desarrolla fetal.
Cuando una hembra es servida, ya sea por monta natural o por inseminación artificial, los músculos uterinos, bajo la influencia de las hormonas Estrógeno y Oxitocina, se contraen rítmicamente para ayudar en el transporte de espermatozoides hacia el Oviducto (Figura 4).
Los oviductos, como su nombre lo indica, conducen los Ovulos, los huevos de la vaca. Los Oviductos son también conocidos como Trompas de Falopio. Los Oviductos presentan varias regiones estructuralmente distintos, al observarlos bajo el microscopio.
La porción más baja, la más cercana al Utero, es llamada Istmo. La coneccion entre el Utero y el Istmo, es llamada Unión Utero-Tubal (UUT). La Unión Utero-Tubal sirve como filtro de espermatozoides anormales y es el reservorio de espermas hábiles (Figura 5).
 Figura 4. Las constracciones uterinas ayudan en el transporte de los espermatozoides.
Figura 4. Las constracciones uterinas ayudan en el transporte de los espermatozoides.
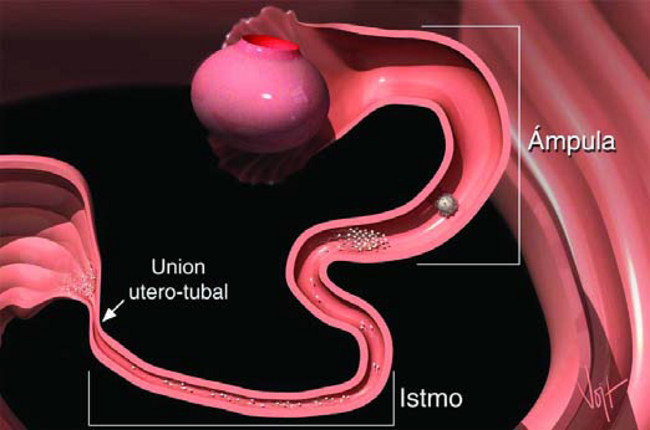
Las investigaciones han sugerido que cuando los espermatozoides llegan al Istmo, estos se adhieren a las paredes. Durante este periodo de adherencia, ocurren muchos cambios fisiológicos a las paredes espermáticas, los cuales son esenciales para que los espermas puedan fertilizar el óvulo. Estos cambios son colectivamente llamados Capacitación, y son aparentemente regulados por esta importante adherencia a las paredes del Istmo. Tarda aproximadamente cinco a seis horas, a partir del momento de la inseminación, para que en el Istmo haya una población espermática capacitada para ejercer la fertilización.
La porción más alta del Oviducto, cercana al Ovario, es llamada Ampula. El diámetro interno del Ampula, adecuando al paso del Ovulo, es mayor que el del Istmo. Es en este segmento del Oviducto donde ocurre la fertilización. Se cree que una señal química, realizada al momento de la ovulación, es la que estimula la liberación de los espermatozoides de las paredes del Istmo, permitiéndoles continuar se viaje al sitio de la fertilización en el Ampula.
La estructura en forma de embudo al final del Oviducto, llamado Infundíbulo, rodea los ovarios y cosecha los huevos, evitando que éstos caigan a la cavidad abdominal (Figura 6).
Estructuras vellosas sobre el infundíbulo y dentro del Ampula, se mueven rítmicamente para transportar el Ovulo y su masa de células Cúmulos, a través del Oviducto al sitio de la fertilización

Figura 6. El Infundibulo atrapa el huevo ala caer del Ovario y lo conduce al Oviducto.

Figura 7. El Ovulo, con su masa de células Cumulus, es transportado al Oviducto por vellosidades.
Los Ovarios son los órganos principales del aparato reproductor femenino. Tienen dos funciones: la producción de Ovulos y la producción de hormonas, principalmente Estrógenos y Progesterona, durante los distintos estadíos del ciclo estral. En la superficie del Ovario se pueden encontrar dos estructuras diferentes: Folículos y Cuerpo Luteo.
Los Folículos son estructuras llenos de fluidos, que contienen los óvulos en desarrollo (Figura 8). Usualmente se pueden encontrar varios Folículos en cada Ovario, que varían en tamaño desde apenas visibles, hasta 20 mm en diámetro. El folículo más grande sobre el Ovario es considerado «el dominante», y es el que probablemente ovule cuando el animal entre en celo. Con el tiempo, más del 95% de los otros Folículos entran en regresión y mueren sin ovular, siendo reemplazados por una nueva generación de Folículos en crecimiento.
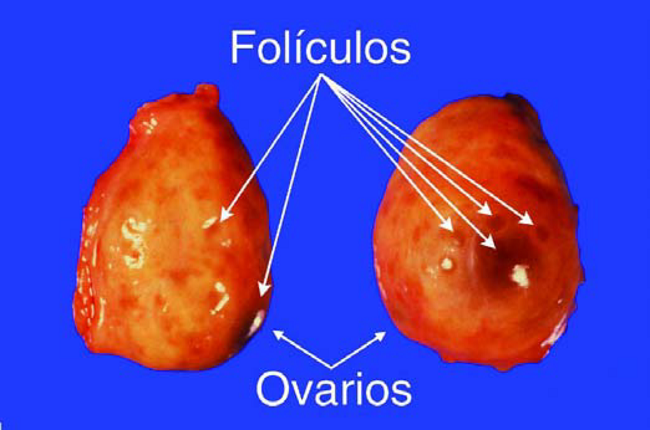
Figuras 8. Folículos sobre los Ovarios.
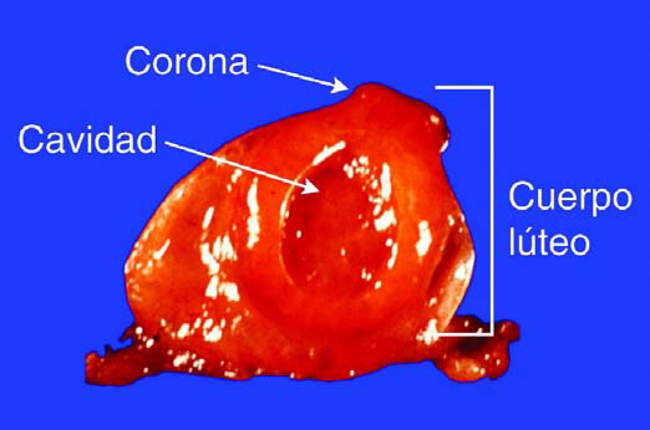
Figura 9. Corte Transversal del Ovario con Cuerpo Lúteo
La otra estructura que se encuentra en la superficie del Ovario es el Cuerpo Luteo (CL). El CL crece sobre el sitio de la ovulación del celo anterior (Figura 9). A menos que haya habido más de una ovalación, se debe hallar solo un CL en uno de los Ovarios.
El CL normalmente tendrá una corona sobre su estructura, lo cual facilita su identificación durante la palpación rectal. El CL también puede tener una cavidad llena de fluidos, pero una pared más gruesa, por lo tanto tendrá una textura más tosca al tacto. El CL en latín significa «cuerpo amarillo.» Aunque en su superficie, esta estructura tiene apariencia oscura, un corte transversal revela un amarillo rojizo en su interior.
EL CICLO ESTRAL
Con el tiempo, ocurren muchos cambios en el aparato reproductor, en respuesta a distintos niveles de hormonas. En una hembra no gestante, estos cambios ocurren cada 18 a 21 días. Esta periodicidad se llama ciclo Estral (Figura 10). Discutamos cómo funciona el ciclo estral, empezando con una vaca en celo al día cero. Si miramos al aparato reproductor, vemos que están sucediendo varias cosas. Un Ovario tendrá un Folículo grande, talvez de 15 a 20 mm de diámetro.
Este folículo contiene un Ovulo maduro, listo para ovular. Las células dentro del Folículo están produciendo la hormona Estrógeno (Figura 11).
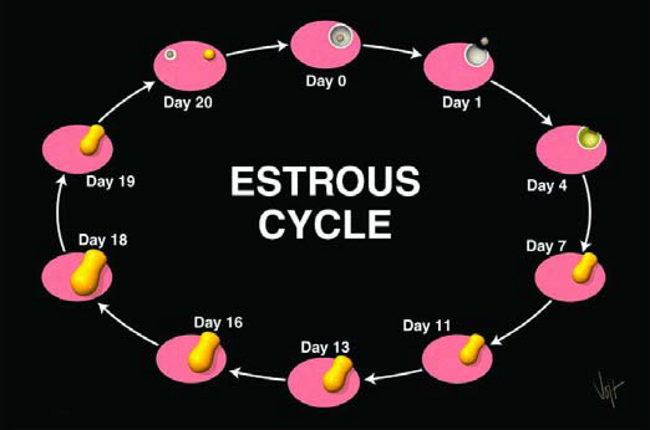
Figura 10. El Ciclo Estral bovino
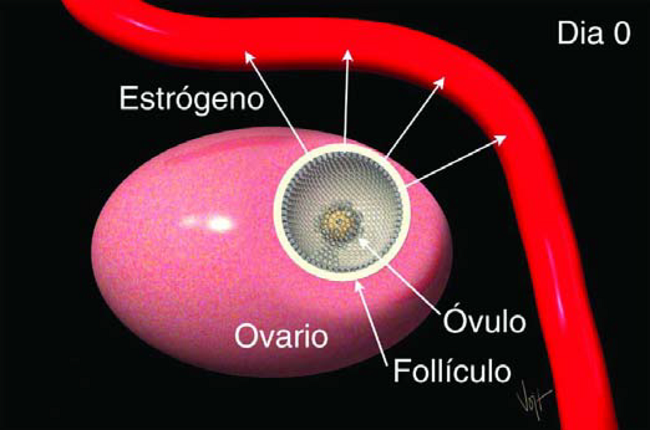 Figura 11. El Estrógeno del folículo en crecimiento es transportado a todo el cuerpo en la corrinte sanguinea
Figura 11. El Estrógeno del folículo en crecimiento es transportado a todo el cuerpo en la corrinte sanguinea
El Estrógeno es transportada por la sangre a todas partes del cuerpo, causando que otros órganos reaccionen de distintas maneras. Hace que el Utero sea más sensible a estímulos, y ayuda en el transporte de espermatozoides después de la inseminación.
Hace que la Cervix secrete un moco viscoso que fluye y lubrica la Vagina. El Estrógeno también es responsable de los síntomas externos del celo, incluyendo una Vulva rojiza y ligeramente inflamada, permitiendo que otras vacas la monten, dejen de comer, mugir frecuentemente y mantener erectas las orejas.
Estos son solo unos cuantos de los muchos síntomas externos del celo.
En el día 1 el folículo se rompe, u ovula, permitiendo la salida del óvulo al Infundíbulo que lo espera (Figura 12). La producción de estrógenos cesa varias horas antes de la ovulación, causando que la vaca no muestre más síntomas de celo.
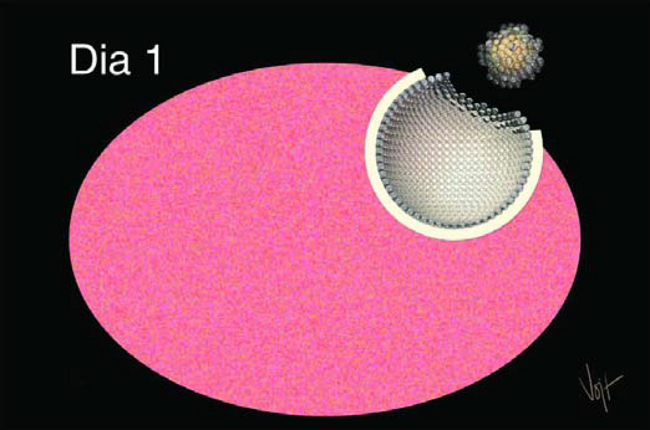 Figura 12. En el sitio de la ovulación crecen células luteicas
Figura 12. En el sitio de la ovulación crecen células luteicas
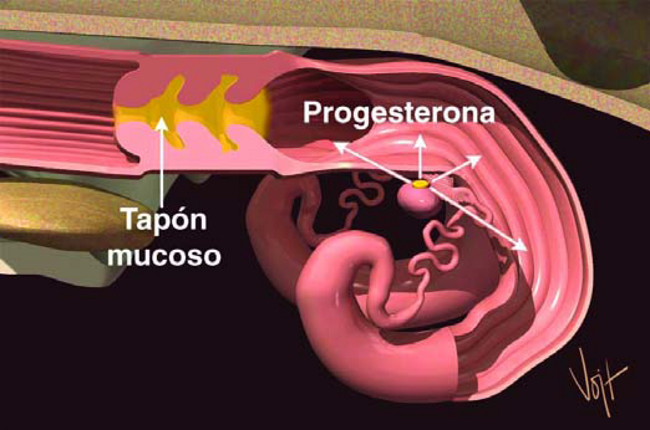 Figura 13. La Progesterona del Cuerpo Lúteo prepara el Útero para la gestación.
Figura 13. La Progesterona del Cuerpo Lúteo prepara el Útero para la gestación.
Después de la ovulación, un nuevo tipo de células, llamadas Células Lutéicas, crecen en el sitio donde estuvo el Folículo.
Durante los próximos cinco o seis días, estas células crecen rápidamente para formar el Cuerpo Luteo (CL). El Cuerpo Luteo produce otra hormona, la Progesterona. La Progesterona prepara al Utero para la gestación (Figura 13). Bajo la influencia de la progesterona, el Utero produce una sustancia nutritiva para el embrión llamada leche uterina. Al mismo tiempo, la progesterona causa que se forme un tapón Muscoso en la Cervix, el cual evita que entren bacterias o virus al Utero.
La progesterona también evita que el animal vuelva al celo al inhibir la liberación de gonadotropinas de la glándula Pituitaria en el cerebro (Figura 14). Existen dos gonadotropinas que la glándula Pituitaria produce, almacena y libera. La primera es la hormona Folícule Estimulante (FSH). Tal como su nombre lo indica, esta hormona estimula el rápido crecimiento de folículos pequeños. La hormona Luteinizante (LH) es la segunda hormona gonadotrópica. Además de ayudar a la producción de Progesterona por el CL, la LH también puede estimular la pro- ducción de Estrógeno por los folículos grandes. Altos niveles de Estrógeno pueden traer al animal de regreso al celo, y complicar la vida del embrión si esta vaca estuviera gestante. Por lo tanto, la regulación que ejerce la Progesterona sobre la producción de FSH y LH es un aspecto crítico sobre el mantenimiento de la
Preñez Por otra parte, si el animal no habia sido inseminada es deseable que vuelva al celo. Los días 16 a 18 del ciclo estral se conocen como » el periodo de reconocimiento materno,» Durante este periodo, el Utero busca la presencia de un embrión en crecimiento.
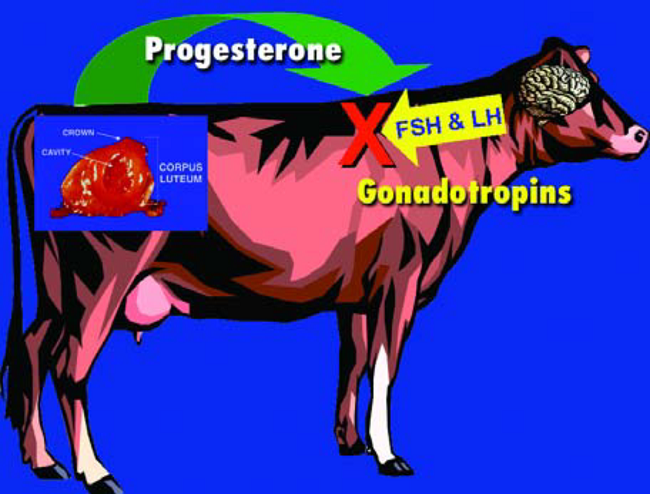 Figura 14. La Progesterona inhibe la liberación de FSH y LH
Figura 14. La Progesterona inhibe la liberación de FSH y LH
Si no se detectara un embrión, el útero inicia la producción de otra hormona, la Prostaglandina. Esta hormona destruye el cuerpo Luteo (Figura 15). Cuando se destruye el CL, cesa la producción de progesterona y la glándula Pituitaria empieza a aumentar la secreción de gonadotropinas. Altos niveles de LH estimulan al folículo dominante a producir Estrógeno y traer al animal de regreso al celo (Figura 16).
 Figura 15. La Prostaglandina destruye al Cuerpo Lúteo
Figura 15. La Prostaglandina destruye al Cuerpo Lúteo
 Figura 16. La FSH estimula el crecimiento de folículos pequeños, mientras que la LH estimula la producción de Progesterona por el CL y la producción de Estrógeno por folículos dominantes.
Figura 16. La FSH estimula el crecimiento de folículos pequeños, mientras que la LH estimula la producción de Progesterona por el CL y la producción de Estrógeno por folículos dominantes.
Con esto se completa un ciclo estral. La periodicidad promedia es de 21 días. El ciclo estral es subdividido en dos fases, dependiendo de la hormona dominante, o en la estructura ovárica presente en cada fase. La fase luteica empieza con la formación del CL, 5 o 6 días después del celo, y termina cuando esta entra en regresión a los 17 o 19 días del ciclo. Durante esta fase, los niveles de Progesterona son altos y los de Estrógeno son bajos.
La otra fase es la folicular. Esta fase inicia cuando el CL de un ciclo entra en regresión y termina cuando se forma el CL del ciclo siguiente. Por lo tanto, la fase folicular abarca el período de la presentación de celo (Figura 17). Durante esta fase los niveles de Estrógeno son altos y los de Progesterona son bajos.
Tal como hemos mencionado anteriormente, pueden haber folículos en los Ovarios en cualquier momento del ciclo estral.
Usando tecnología de ultrasonido, las investigaciones han detectado que la aparición de folículos sobre los Ovarios ocurre en «olas.» En un ciclo estral normal de 21 días, un animal puede experimentar 2 o 3 olas de crecimiento folicular (Figura 18). El inicio de cada ola se caracteriza por un pequeño incremento de FSH, seguido por el rápido crecimiento de varios folículos. De esta ola folicular, un folículo es escogido para crecer más que los otros. Este folículo «Dominante» tiene la habilidad de restringir el crecimiento de todos los otros folículos en los ovarios.
Los folículos dominantes solo duran de 3 a 6 días, que es cuando mueren y entran en regresión, u ovulan. En consecuencia, la desaparición del folículo dominante coincide con la formación de la siguiente ola, del cual saldrá otro folículo dominante.
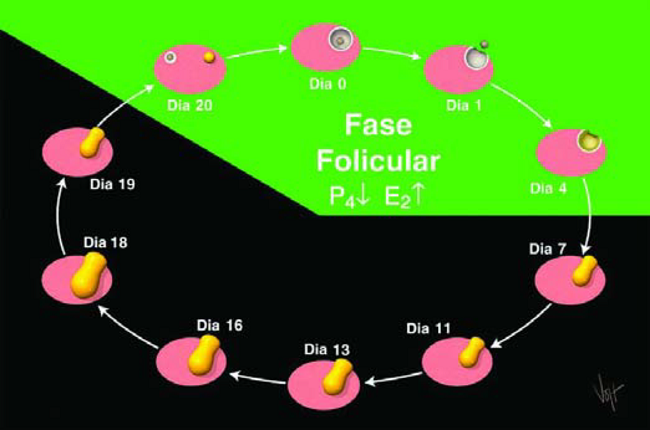 Figura 17. El Ciclo Estral se divide en dos fases.
Figura 17. El Ciclo Estral se divide en dos fases.
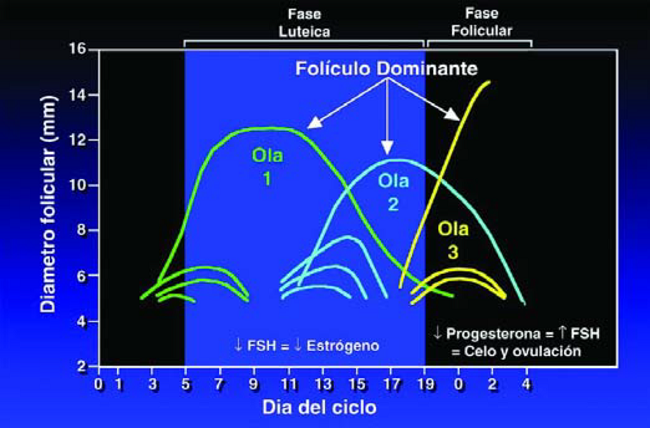 Figura 18. Hay crecimiento folicular en todo el Ciclo Estral, pero los niveles de Estrógeno solo suben durante la fase folicular.
Figura 18. Hay crecimiento folicular en todo el Ciclo Estral, pero los niveles de Estrógeno solo suben durante la fase folicular.
Aunque sea normal tener crecimiento folicular durante todo el ciclo estral, los bajos niveles de LH durante la fase lutéica, evitan que estos filículos produzcan altos niveles de Estrógeno, lo cual traería al animal de regreso al celo. Solamente el folículo dominante presente al momento de la regresión del CL, cuando los niveles de Progesterona son bajos, puede producir suficiente Estrógeno para traer al animal al celo y continuar hasta la ovulación.
DESARROLLO DEL EMBRION Y DEL FETO
Durante los primeros 4 o 5 días el embrión viaja por el Oviducto hacia el Utero. Una vez que el embrión llegue al Utero, será rodeado de fluidos uterinos y seguirá su crecimiento.
Mientras este embrión esté flotando libremente en el útero, varias membranas se están formando, incluídos el Amnion, el Corion y el Alantoides. En su conjunto, estas membranas son conocidas como la Placenta.
Idealmente, cuando llegue el periodo de reconocimiento materno, entre los días 16 a 18, el feto y la Placenta en crecimiento habrán producido suficientes cantidades de señal química necesaria para mantener la gestación. Esta señal inhibe el efecto de la Prostaglandina sobre el CL (Figura 19). El CL entonces se mantiene intacto y continúa produciendo Progesterona, hormona vital para mantener la gestación.
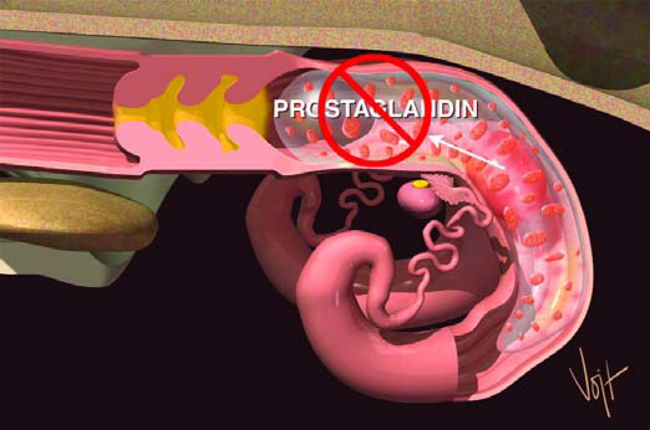 Figura 19. El feto inhibe la producción de prostaglandina.
Figura 19. El feto inhibe la producción de prostaglandina.
Alrededor de los 30 días de gestación, la placenta empieza a adherirse al Utero en varios puntos. El sitio de adherencia al lado de la placenta se llama Cotiledón, mientras que al lado del Utero se desarrollan Carúnculas. La unión entre Cotiledones y Carunculas es como una mano en un guante. Esto aumenta grandemente la superficie de adherencia en el sitio de la unión, facilitando el intercambio de nutrientes y desechos entre la cría y la madre, por venas y arterias que viajan hacia y a través del cordón umbilical.
Al momento del parto, los músculos empiezan a contraerse y expulsan al ternero y sus membranas a través de la Cervix y la Vagina ya dilatadas. Varias hormonas, incluyendo Progesterona, Estrógenos, Prolactina, Relaxina y Corticoides, producidos por la madre, el feto y la placenta, se conjugan para realizar este evento (Figura 20). El hecho de dar a luz en un sitio limpio y el cuido de la vaca después de un parto, ayudan a prevenir problemas reproductivos.
Mientras más se familiarice con la anatomía y fisiología de la vaca, mejor será el trabajo de inseminación artificial que se realice. Un claro entendimiento de la interacción de las hormonas que afectan el ciclo estral, te brinda una mayor comprensión del porque se presentan los variados síntomas de celo, como se mantiene la preñez, y que se debe hacer si las vacas no ciclan normalmente.
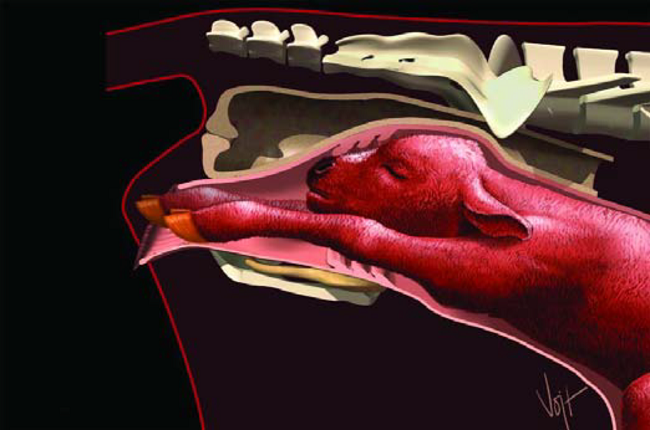 Figura 20. Producto Final
Figura 20. Producto Final











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
-

 Artículos1 año atrás
Artículos1 año atrásGuía de Manejo de Semen Bovino
-

 Artículos1 año atrás
Artículos1 año atrásDetección de celos en vacas: un punto crítico para el éxito reproductivo
-

 Noticias Nacionales11 meses atrás
Noticias Nacionales11 meses atrásConsumo de bebidas: ¿cuál es la categoría que más crece en 2025 y por qué?
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásGloria lanza programa “Productividad Ganadero” a favor de la producción de leche en el Perú
-

 cursos on line1 año atrás
cursos on line1 año atrásCurso Virtual: Actualización en Sanidad Bovina – Control, Inmunología y Manejo de Vacunas
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásAgrovet Market lanza nueva línea de antibióticos en suspensión para el tratamiento de la mastitis
-

 Noticias Nacionales1 año atrás
Noticias Nacionales1 año atrásGloria: la primera marca peruana en el Kantar Creative Effectiveness Awards 2025
-

 Cursos A Distancia7 meses atrás
Cursos A Distancia7 meses atrásCurso Virtual: Formulación de Raciones para Ganado Lechero con software especializado NASEM